

Faucons



Le mot “Faucon” a pour origine le mot latin “falco”, dérivé lui-même de “falx” la faux ou “falcicula” la faucille : les ailes des faucons ayant la forme d’une lame de “faux” – “falciforme” (et non pas, comme on le lit parfois, parce que les serres ou le bec sont “falciformes”, puisque tous les rapaces et bien d’autres espèces, présentent ces mêmes particularités non spécifiques aux faucons).
Étonnement, les faucons longtemps classés avec les autres rapaces – ils ont même donné leur nom aux groupes des falconiformes et falconidés – voisinent maintenant avec les Psittaciformes (perroquets et perruches au sens large), comme l’ont montré les études génétiques récentes. Perruches et perroquets partagent il est vrai avec les faucons nombre de particularités anatomiques : tête ronde, bec crochu, ailes falciformes, vol battu direct et rapide, et pour certaines espèces, narines rondes pourvues d’un “frelon” – sorte de petit bouton situé au centre de chaque narine ( 1) 
(1) Ce qui ferait perdre de sa pertinence à la théorie – colportée un peu partout sur internet – selon laquelle “le frelon” aurait été sélectionné au fil de l’évolution pour permettre aux faucons de respirer quand ils plongent sur leurs proies à grande vitesse ? Or tous les faucons ne piquent pas à grande vitesse, mais tous, et nombre d’autres espèces génétiquement apparentées ou non, les caracaras, certains psittacidés, les buses de Harris, l’épervier d’Europe, par exemple, ont des narines pourvues d’un “frelon”. Ce qui amènerait à penser , si on s’en tient à cette explication, que le faucon pèlerin spécialiste de la chasse en piqué à des vitesses pouvant dépasser 300-350 km/h, donc chez qui le frelon aurait l’utilité qu’on lui prête, serait l’ancêtre de toutes les espèces pourvues d’un “frelon” : faucons, caracaras, psittacidés, buses de Harris et épervier, lesquelles auraient conservé cet attribut sans utilité pour elles ? Cela semble peu vraisemblable pour une si grande diversité d’espèces, d’autant que faucons et accipitridés ne sont pas du tout apparentés au plan génétique. De plus, il est généralement admis que l’évolution se fait du “généraliste” vers le “spécialiste” et non pas l’inverse : les analyses montrent en effet, qu’au sein d’un groupe d’espèces génétiquement proches, ce sont les plus spécialisées les plus récentes (étude génétique). Et pourquoi l’aigle royal capable de piquer à près de 300 km/h, du fait de sa masses imposante (4 à 7 kg), donc plus vite que tous les faucons – pèlerin exclu – n’en est-il pas “équipé” ? Par ailleurs il est à noter que tous les oiseaux sont “dotés” de sacs aériens – “réservoirs” d’air améliorant et régulant la respiration et la température interne (et non pas la sudation, car les oiseaux n’ont pas de glandes sudoripares)- qui permettent en outre aux oiseaux de survivre de longues périodes dans l’air raréfié des hautes altitudes. Or les piqués de chasse à grande vitesse ne durent que quelques secondes, une quinzaine tout au plus pour les plus longs. Pendant un laps de temps aussi court, un faucon n’aurait aucun mal à résister à une ventilation perturbée. En conclusion, il semble bien que l’hypothèse, certes séduisante, selon laquelle le “frelon” jouerait le rôle de “décompresseur” à l’entrée des narines lors des piqués à grandes vitesses, ne résiste pas à l’analyse, d’ailleurs certains lézards et serpents aussi bien que les tyrannosaures avant eux, qui n’ont pourtant pas la réputation de piquer à grande vitesse, en sont également pourvus. Le rôle du “frelon”, déjà présent chez certains dinosaures, reste donc encore un mystère. À moins que ? Nouvelle hypothèse ?
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
L’Europe compte 10 espèces de faucons : Gerfaut, Sacre, Pèlerin, Lanier, Émerillon, Crécerelle, Crécerellette, Hobereau, Faucon d’Éléonore et Kobez, dont 4 nichent en France : Pèlerin, Hobereau, Crécerelle, Crécerellette. Les faucons sont suffisamment proches génétiquement pour être tous “hybridables” en reproduction captive. En nature, des cas d’hybridation naturelle ont été observés entre pèlerin et lanier en Italie, pèlerin et faucon de prairie en Amérique du Nord, entre sacre et gerfaut dans l’Altaï en Asie centrale. Les caractéristiques anatomiques et comportementales des faucons sont très semblables, de même que la forme des ailes et le vol battu rectiligne (voir début de page). Ce qui les distingue entre eux, c’est la corpulence, la couleur du plumage, les caractéristiques de la voilure – surface des ailes et de la queue, les techniques de chasse, la longueur des serres et le régime alimentaire. Une queue longue, une surface alaire importante, des plumes portantes relativement souples, sont des caractéristiques qui permettent une plus grande “manoeuvrabilité, favorisant la captures de proies terrestres. Au contraire, une queue courte, un plumage rigide, une voilure plus compacte, combinés à une masse relativement importante, facilitent des vitesses d’attaque en piqué nettement supérieures, conduisant à des spécialisations comportementales et des évolutions morphologique mieux adaptées à la captures de proies volantes (Les doigts remarquablement longs du faucon pèlerin).
{kind=link}
LA VUE EST LE SENS LE PLUS DÉVELOPPÉ CHEZ LES OISEAUX
La couleur des yeux :
Les faucons, toutes espèces confondues, sauf une sous espèce de faucon crécerelle du SE africain dont l’iris est blanchâtre – Falco rupicolus ou rupicoloides – ont les yeux noirs (ne pas se laisser abuser par le tour de l’œil dépourvu de plumes qui est de couleur jaune profond chez les adultes). C’est d’ailleurs pour cette raison qu’en fauconnerie orientale les faucons sont nommés les « yeux noirs », au contraire de l’autour et de l’épervier qui sont les « yeux jaunes ».
L’anatomie de yeux :
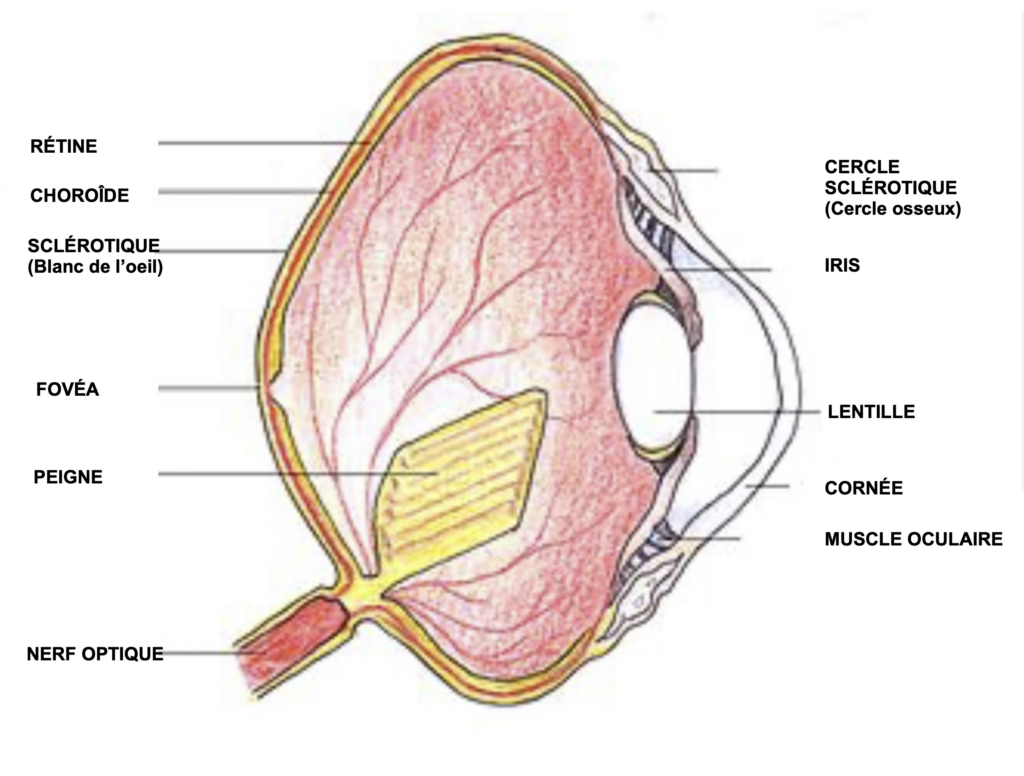
Les caractéristiques anatomiques des yeux des rapaces sont globalement les mêmes que chez les mammifères, mais ils sont ÉNORMES (60 à 70% du volume du crâne). À l’échelle humaine ils auraient la taille d’un gros pamplemousse.
Par ailleurs, ils ne sont pas sphériques mais légèrement aplatis, ce qui leur donne une plus grande surface rétinienne, d’où un plus grand nombre de cellules visuelles disponibles – l’équivalent des pixels pour les appareils photographiques.
La contrepartie de cette grande taille est une faible mobilité, voire pas du tout – rapaces nocturnes par exemple.
Chez les faucons la partie postérieure interne de l’œil, la RÉTINE, est tapissée de cellules sensibles à la lumière – les cellules visuelles ou photorécepteurs. Alors que la rétine de l’homme ne dispose que de 4 sortes de ces cellules :
– 1 en forme de bâtonnet pour la vision en nuances de gris (vision pour les faibles intensités lumineuses et la vision nocturne). Environ 20 000 à 50 000 bâtonets/mm ils sont répartis dans toute la périphérie de la rétine et absents de la fovéa) .
– 3 pour la vision des couleurs rouges, vertes et bleues (peu nombreuses en dehors de la fovéa où ils sont en grand nombre – 150 000 à 180 000 cônes/mm2 ).
La rétine des rapaces compte 6 sortes de cellules visuelles.
– Les bâtonnets et les cônes pour le rouge, le vert et le bleu, comme chez l’homme, mais deux autres types de cellules en cône y sont présentes :
– 1 pour l’ultra violet .
– 1 en forme de double cône pour la régulation de l’ensemble des autres cellules visuelles (peut être sensibles aux longueurs d’ondes radio-électriques – les très grandes longueurs d’ondes au delà de l’infrarouge).
Par ailleurs, au centre de toutes les cellules en « cônes » des rapaces diurnes, à la naissance des cônes, est présente une gouttelette sphérique de graisse colorée de 2-3 microns de diamètre. Cette micro lentille concentre la lumière sur le cône proprement dit, provoquant un accroissement conséquent de l’acuité visuelle (indépendamment de la haute densité des photorécepteurs). De plus leur coloration filtre la lumière en éliminant les franges marginales de la fréquence lumineuse pour ne laisser passer que les photons les plus proches de la couleur dominante, rendant ainsi la perception des couleurs plus intense.
De plus les yeux de tous les rapaces diurnes comme nocturnes, possèdent une couche de cellules réfléchissante juste derrière les cellules visuelles – le tapetum lucidum. Ce “miroir”, en contact direct avec l’extrémité externe des cônes, renvoie la lumière vers l’avant, augmentant ainsi la quantité de lumière qui active le photorécepteurs. Toutes ces différences avec l’anatomie de l’oeil humain permettent aux rapaces d’avoir une acuité visuelle estimée environ 6 à 8 fois supérieure à la nôtre.
De plus la rétine des oiseaux chasseurs , donc les rapaces (mais pas les vautours qui ne sont pas des chasseurs mais des charognards, ni les nocturnes), sont “équipés” de deux fovéas – région de l’œil ou la densité des cellules visuelles est la plus grande (les mammifères n’en disposent que d’une seule).
– La fovéa principale dans l’axe de l’œil, dite “nasale“, est celle dont la densité de cellules – uniquement composée de cellules en cônes- est la plus élevée. C’est avec cette partie de l’œil que les faucons repèrent les proies à très grandes distances. Comme l’axe de l’œil est orienté à environ 40° vers l’avant, cela explique pourquoi les faucons inclinent ou tournent la tête sur le côté pour analyser d’un seul œil, ce qu’ils remarquent à grandes distances.
– La fovéa secondaire, dite fovéa «temporale », est située plus vers l’extérieur de l’oeil. La densité des cellules visuelles est moindre que celle de la fovéa principale, mais permet la vision binoculaire plus efficace la poursuite des proies.
Par ailleurs dans la partie postérieure interne de l’œil se trouve un organe (une sorte de peigne), fortement irrigué dont la fonction n’est pas très claire.

Dimorphisme sexuel.
Chez les rapaces – diurnes et nocturnes, hormis les charognards – le mâle est toujours – plus ou moins – plus petit que la femelle. D’une façon générale plus les proies chassées par une espèce sont rares et/ou difficiles à capturer – spécialement les oiseaux volants – plus l’écart de taille mâle-femelle est important en faveur de la femelle – Épervier, faucon pèlerin, faucon à poitrine orangée, par exemple. Au contraire plus les “proies” sont abondantes et/ou faciles à capturer – rongeurs, reptiles, carcasses, etc… – moins l’écart est perceptible de visu – buses, busards, milans, circaète, vautours, etc… Chez les buses et les busards, par exemple, les proies sont constituées essentiellement de rongeurs – espèces abondantes – la différence de taille mâle/femelle n’est pas ou peu sensible à l’oeil. Chez les vautours – consommateurs de carcasses, « proies faciles à capturer” – l’absence de dimorphisme est encore plus net, nul ou quasi nul, même sur la balance. Le dimorphisme sexuel apparait donc avant tout comme une adaptation permettant aux deux partenaires d’un même couple de ne pas entrer en concurrence alimentaire quand les ressources sont rares ou difficiles d’accès. La femelle chasse de grosses proies alors que le mâle se contente des plus petites.
Mais pourquoi est-ce la femelle la plus grande et non pas l’inverse ?
Chez les oiseaux on distingue les nidifuges des nidicoles. Chez les nidifuges les poussins quittent le nid dès la naissance – c’est le cas des gallinacés, entre autres. Chez les nidicoles, dont font partie les rapaces, les jeunes restent au nid sous la protection des adultes jusqu’à l’émancipation. Ce comportement différent des jeunes conduit à des formes d’élevage différentes, d’où découle une pression de sélection et des évolutions morphologiques différentes. Le cas emblématique est celui du faucon pèlerin.
Chez cette espèce les oeufs (rougeâtres) sont aussi gros que ceux d’une poule (3 à 4 fois plus lourde qu’une femelle de faucon). La femelle couve les 2/3 du temps – au moins – il est donc clair, vue la taille des oeufs, qu’une forte corpulence favorise une incubation plus efficace. Mais passer plus de temps à l’aire à couver, réchauffer, nourrir et protéger les jeunes, augmente les probabilités de confrontations avec des prédateurs. Une forte corpulence, donc une grande force physique, amplifiée par une agressivité inter-spécifique considérable, constitue un atout favorable à la protection de la nichée.
Ainsi, chez le faucon pèlerin le mâle est sensiblement ⅓ plus petit que la femelle, ce qui lui a valu le nom de “tiercelet” en fauconnerie. (Depuis plusieurs décennies ce qualificatif est également attribué à tous les mâles de rapaces même si l’écart de taille mâle/femelle n’est pas aussi marqué que chez le pèlerin. En Franche-Comté le faucon crécerelle est aussi nommé “tiercelet” par confusion avec le pèlerin). Dans l’est de la France le tiercelet – le vrai – pèse de 550 à 600 g, plus rarement jusqu’à 650 g, la femelle – la “forme” – de 900 à 950 g – aux extrêmes 800 et 1200 g plus rarement. Certaines femelles sont donc 2 fois plus lourdes que leur partenaire.
Pour résumer de façon amusante (peut-être) : pendant que “Madame” reste à “la maison” à s’occuper de la marmaille, Monsieur est «au boulot ». La famille Faucon semble très traditionnelle, de type “patriarcal”, sauf qu’en réalité c’est tout le contraire. Par sa grande taille, sa force et son agressivité inter-spécifique considérable, la femelle est la “dominante” au sein du couple, c’est elle “la patronne”.
Le FAUCON PÈLERIN : Vocalisationsvocalisations.
Le chant – ou cri long – du mâle est plus clair que celui de la femelle, il est émis surtout à la période des pariades de février à mai. Lors des parades nuptiale – février-mars – plus spécialement avant la ponte, quand mâle et femelle se retrouvent sur l’aire, face à face, bec pointé vers le sol, corps incliné en avant, queue plus ou moins verticale – sortes de «salutations » (voir répertoire comportemental) – ils émettent alors des cris brefs “claquants”, qui pourraient être transcrits par «tsick », « tiock », « iiitchop » (Dans cette dernière séquence extraite du film de JP Macchioni “La falaise aux pèlerins“, s’intercale le “chant”, ou cri long de la femelle, plus grave que celui du mâle). Des cris semblables, associés à une attitude similaire – tête basse, queue pointée en l’air, qu’on pourrait considérer comme l’attitude d’agressivité intra-spécifique caractéristique – sont également émis lorsqu’un faucon étranger pénètre sur le territoire de reproduction d’un couple cantonné. Ce type de comportement territorial est surtout développé au printemps, un peu moins à l’automne lors des passages de faucons migrateurs. À l’occasion des nourrissages à l’aire l’adulte qui distribue les becquées – le plus souvent la femelle – émet des “chocks’ insistants qui déclenchent l’ouverture du bec des jeunes poussins, ou la sollicitation des plus grands (Jeunes de 10-15 jours). Quand les fauconneaux atteignent l’âge de 3 semaines, environ, leurs cris “éraillés” et lancinants sont caractéristiques. Il sont émis par les jeunes lors des nourrissages à l’aire, ou à la vue d’un adulte en vol jusqu’à leur émancipation, 1 à 1,5 mois après le premier vol. Le cri d’alarme – ici la femelle, voix plus grave que celle du “tiercelet” – est émis quand un adulte ou un jeune bien développé perçoit une menace potentielle.
Jeune faucon pèlerin jurassien mâle mangeant une grive (Pour éviter toute perturbation Alexis Leclerc utilise une camera automatique, prépositionnée.
Pourquoi le faucon pèlerin est-il l’animal le plus rapide au monde ?
LES ATTAQUES DE CHASSE.
Les attaques du faucon pèlerin sont initiées à partir de 2 points de départ possibles
- Soit depuis un poste d’observation élevé, au sommet d’une falaise ou d’un arbre.
- Soit à partir d’un “vol amont”, le faucon cerclant très haut dans le ciel, parfois hors de vue sans jumelles (2-3 km ). Ce “vol d’amont” à grande hauteur permet au faucon de repérer – d’aveuier (terme de fauconnerie) – des proies potentielles à de très grandes distances – jusqu’à plus de 10 km. Mais aussi en augmentant la pente du piqué d’en accroître la vitesse – fréquemment plus de 250 voire plus de 300 km/h. Les études en soufflerie et les calculs associés du bio-physicien américain, Vance A. Tucker, tendent à montrer que lors qu’un piqué vertical, un faucon de 1kg, pourrait théoriquement dépasser la vitesse de 410km/h. Cette vitesse maximale théorique nécessiterait une chute verticale d’au moins 1200m, circonstances qui ne se produisent jamais en Nature. La vitesse maxima mesurée de 394 km/h l’a été lors d’un piqué vertical “à blanc”, car en chasse les piqués verticaux jusqu’à la proie ne sont pratiquement jamais observés, tout simplement parce que la proie vole et s’éloigne de la verticale du faucon ce qui “aplatit” l’angle d’attaque, réduisant ainsi la vitesse.
- Une fois déclenchée, l’attaque comporte deux parties : un vol de placement et un piqué terminal aboutissant (ou pas) à la capture de la proie.
- Le vol de placement est un vol battu énergique et soutenu sans phase de plané (ou très peu). Il se distingue ainsi du vol de migration ou de déplacement ordinaire constitué d’une alternance de quelques battements d’ailes et de planés. Il peut se dérouler sur une trajectoire montante formant une large courbe sans rapport apparent avec la position de la proie, mais le plus souvent il se développe en direction de celle-ci selon une trajectoire rectiligne horizontale ou montante.
- Le piqué terminal. Quand le vol de placement s’incline vers la proie qui vole plus bas, c’est l’amorce du piqué terminal. Lors de ce piqué le faucon alterne battements d’ailes très rapides et étriqués, presque “mécaniques”, avec des phases ailes fermées. Il est suivi du piqué terminal proprement dit, ailes totalement collées au corps avec, éventuellement de loin en loin, l’écartement latéral des bouts d’ailes (sans doute pour ajuster la vitesse et la trajectoire du piqué ?) Voir la video). Le piqué aboutit au “contact” avec la proie, qui peut être “liée”, c’est à dire prise et tenue fermement avec les serres (la proie peut rester vivante après la capture), soit “buffetée”, c’est à dire frappée au passage avez les serres. Dans ce cas la proie tuée sur le coup tombe vers le sol. Le faucon la récupère dans un deuxième temps en la “liant” après une “ressource” (changement de trajectoire brutal vers le haut ou latéralement) et un piqué secondaire qui aboutit à la capture définitive.
- On a pu lire dans un article scientifique que la trajectoire terminale du faucon n’était pas rectiligne dans un plan horizontal, mais dessinait une courbe. Cette hypothèse était fondée sur des mesures faites en soufflerie qui montrent des modifications de vitesses importantes selon le positionnement de la tête, et sur l’hypothèse que le faucon en piqué utilise sa vision monoculaire et non pas binoculaire. De sorte que, en effet si pendant le piqué le faucon tournait la tête latéralement pour conserver l’image de la proie sur sa fovéa principale (Celle dont l’acuité est la plus grande, la fovéa étant située dans l’axe de l’œil, et orientée à 40° vers l’avant du fait de la configuration anatomique de la tête, vision monoculaire), il réduirait son coefficient aérodynamique et perdrait de la vitesse. Or pour conserver la tête dans l’axe tout en gardant l’image de la proie au centre de l’oeil, le faucon ne devrait pas piquer en direction de celle-ci mais à côté, ce qui se traduirait effectivement par une trajectoire courbe.
- En plus de 60 ans d’observations, hormis quand la proie se déplace latéralement, cas le plus fréquent, où le piqué s’infléchit selon une trajectoire courbe, dite “courbe du chien” ou “courbe de poursuite“, je n’ai jamais constaté de courbure particulière des trajectoires d’attaque quand la proie se déplace en ligne droite en venant ou en s’éloignant du faucon. Selon moi la fovea principale, la plus performante, est utilisée seulement pour repérer et identifier les proies à grande distance (vision monoculaire), voire aussi, pourquoi pas, pendant la phase de placement, phase pendant laquelle une perte de vitesse due aux mouvements de la tête a peu d’incidence sur la vitesse (évaluée entre 80 et 90 km/h). Mais pour le piqué terminal le faucon garde la tête fixe, dans l’axe du corps, la proie étant alors suivie en vision binoculaire grâce aux deux foveas secondaires – temporales – dont l’évolution les a justement sélectionnées pour ça, la poursuite des proies.
- Au moment du contact avec la proie, qu’elle soit “liée” ou “buffetée”, l’attitude du faucon est très stéréotypée. C’est un comportement inné qui se manifeste déjà chez les jeunes faucons en premier duvet. Le faucon redresse le corps, tête en arrière, ailes à moitié ouvertes en arrière des épaules (comme celles des anges), en même temps qu’il projette les serres en avant (Ce que font en fait tous les rapaces au moment d’une capture) avec la nuance, pour le faucon pèlerin, que cette séquence se produit à si grande vitesse qu’elle est longtemps restée inaperçue sans les moyens techniques actuels (video à grande vitesse). À l’oeil nu on a l’impression que le faucon passe à côté de la proie sans la toucher et sans modifier son attitude, ce qui a fait croire aux fauconniers pendant longtemps que la proie n’était pas frappée, “buffetée”, avec les serres mais avec le thorax (Voir la video).
- Le taux de réussite des attaques du faucon pèlerin est extrêmement variable selon la région, le type de proies attaquées, la période de l’année et les individus. Il est admis que le taux de réussite moyen se situe autour d’une attaque réussie pour 4 à 6 attaques infructueuses. Dans le Jura, le taux est plus élevé, une capture pour 10 attaques. J’ai même observé en période de migration printanière 22 attaques manquées avant que la femelle réalise une prise à sa 23eme tentative. On est donc bien loin de l’oiseau mythique qui prend à chaque fois.
- La taille des proies elle aussi varie, comme le taux de réussite, selon la région. Dans le Jura, leurs tailles s’étalent, grosso modo, de celle du pinson à celle du pigeon ramier, avec une fréquence maximale pour les oiseaux de tailles intermédiaires – étourneaux, merles, grives et geais. Dans d’autres régions – falaises côtières du pays de Caux par exemple – la grande majorité des proies est constituée de pigeons domestiques retournés à l’état sauvage – pigeon feral – nicheurs abondants dans les falaises du littoral de la Manche. En résumé il n’est pas raisonnable de vouloir définir précisément « LE » régime alimentaire du faucon pèlerin. On a juste le droit de dire que « le faucon pèlerin est un chasseur d’oiseaux capturés en vol (ou même de toutes sortes d’animaux volants – insectes, chauves souris, les oiseaux représentant bien entendu l’immense majorité des captures).
Quelques informations concernant les faucons en général et plus spécialement le faucon pèlerin, articles et videos :
- Faucons caractéristiques communes.
- Le faucon pèlerin biologie (Monographie).
- Incubation et elevage. (Video)
- Zoom sur le faucon pèlerin. Article en français.
- Répertoire comportemental du faucon pèlerin. Article en français.
- Comportement particulier : l’aide à l’élevage. Article en français.
- Faucon pèlerin chronométré à 389 km/h. Vidéo anglo-américain.
- Le faucon pèlerin jurassien. Vidéo en français.
- Évolution de la population jurassienne de faucon pèlerin. Communication Varsovie 2008. Anglais.
- Évolution de la population française du faucon pèlerin jurassien. Communication Varsovie 2008. Anglais.
- Le faucon pèlerin en Normandie.
- Évolution de la population du faucon pèlerin. Analyses statistiques. Communication Budapest 2017. Anglais.
- Faucon pèlerin nord américain. Video.
- Faucon pèlerin vs grand-duc, cohabitation impossible (sauf cas exceptionnels) article.
- Génétique des 3 sous espèces nord américaines.
- BILAN PROSPECTION 2024.